Unicellular organisms
Unicellular organisms have cellular communication mechanism as well. One hypothesis to explain why complex organisms came about with a delay is due to the time needed to develop cellular signalling mechanism.
- Many bacteria can release and sense chemical signals
- Signalling b/w yeast cells prepares them to mate
- Signalling b/w ameboid cells can cause them to aggregate e.g. Dictyostellium cells come together to form a fruiting body in order to have spores spreading to other areas.
Multicellular organisms
mammals, flies and worms have similar communication pathways
Sending signals
- signals are sent out in different forms: proteins, small peptides, nucleotides, dissolved gases, etc.
- signals are sent out in different ways: by exocytosis, diffusion, displaying on cell surface
Receiving signals
- signals can bind to surface receptor proteins if they are hydrophilic (cannot pass the hydrophobic membrane)
- signals can be transported on carrier proteins and then bind a intracellular receptor if they are hydrophobic
- more combinations are possible
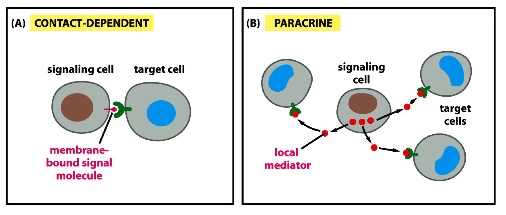
Short Distance Signalling
1) contact-dependent
- signalling molecules on the signalling cell must be directly in contact with the receptor on the target cell (e.g. Notch signal)
- signals are retained on the cell surface
2) paracrine
- signals are released and diffused away but only act locally
- signals are restricted due to:
-
- internalization by neighbouring cells
- signal instablized or destroyed by extraceullar enzymes
- signals bind to extracellular matrix molecules
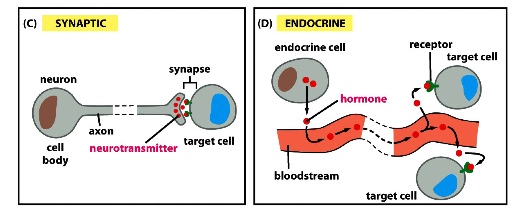
Long Distance Signalling
1) synaptic signalling
in nervous system, neurons extend axons to contact distant target cells — the long range is due to the long length of the axons
but the released signalling molecules only act locally at target
2) endocrine signalling
hormones are released into the bloodstream and carried to target far away
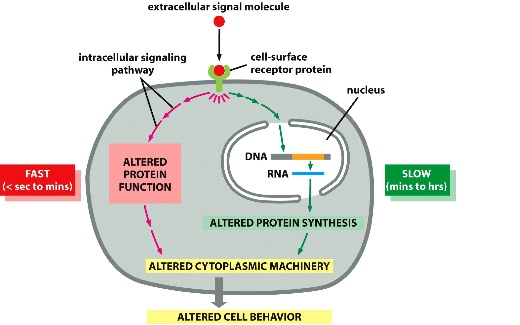
Cells have signal transduction pathways to respond to extracellular signals
Definitions
Signal transduction: conversion of extracellular signal to intracellular signal (or in general, any type of signal converted to another type)
Effector: a downstream molecule in a signal transduction pathway —> upstream molecuels have effects on them
Between extracellular signal and the final effector proteins, there are small intracellular signalling molecules or large intracellular signalling molecules. —> both bind to their downstream molecule and change their conformation in some way.
A closer look at the components of signal transduction pathways
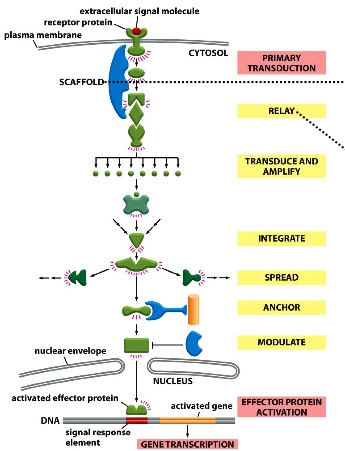
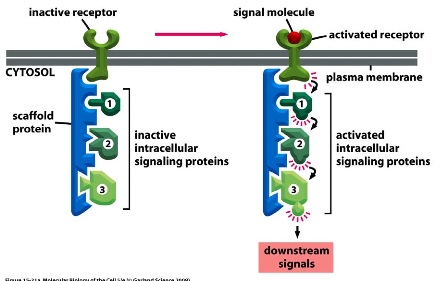
- Scafold protein: draws together multiple signalling components in order to increase efficiency and specificity
- Relay: sends signal downstream in the same basic form —> no alteration, no transduction (e.g.protein-protein interactions)
- Transduce and amplify: signals are converted into different forms. If there’s a protein, the one protein can convert many downstream targets and thus amplifying them
- Integrate: coordinate and combine signals from different sources —> when more than one signal is required to activate downstream targets (coincidence detector makes sure that all the requirements are met)
- Spread: sends signals to other pathways
- Anchor: restricts signals to certain part of the cell —> localizing signalling event
- Modulate: inhibits signals to restrict the intensity of signalling (= down-regulation)
Many components of signalling pathways act like binary switches. In order to turn the switch on, it also need to be turned off now and then. Similarly, with signalling pathways, it’s important to activate signalling molecules, but it’s also important to inhibit signalling molecules.
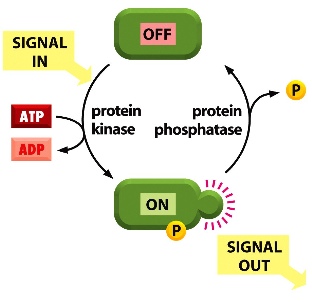
Phosphorylation
- One of the most important signalling process in humans.
- Phosphate groups are added by protein kinases, and removed by protein phosphatases.
- There are serine/threonine kinases and tyrosine kinases: they residues have -OH groups on them where phosphate groups can be added on
- signal comes in —> protein kinase uses the phosphate group on ATP and add it onto the target protein —> the conformation of the phosphrylated protein is changed —> sends a signal to the downstream targets —> when the signal is not in use, protein phosphatase removes the phosphate group and turns the protein off
GTP or GDP binding
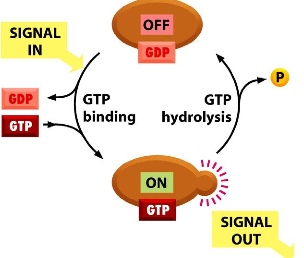
- There are large trimeric and small monomeric types
- They have low GTPase activity by themselves, so they needs extra helpers: GAPs and GEFs
-
- GAPs = GTPase activating preteens increase GTP hydrolysis
- GEFs = Guanine nucleotide exchange factors remove GDP and allow GTP to bind
- signal comes in —> GEF removes GDP, allowing new GTP to bind —> conformation of the protein is changed —> sends a signal to the downstream targets —> when the signal is not in use, GAP promotes GTP hydrolysis and turns the protein
Protein interaction
Proteins often contain one or more interaction domains. A single polypeptide chain can have multiple folded semi-automic domains that have different properties and binding characteristics.(e.g. SH2, PTB, PH, SH3)
During evolution, domains can be added or switched to alter interactions and re-wire signalling pathways.
How is signalling specificity achieved?
1) Have short range signalling so that the signals can act directly to the local target
2) Synaptic signalling: neurons make connection only to its target cells. Once the links are made, specificity can be maintained.
3) Endocrine signalling: the signalling molecules can only bind to specific receptors on the target cells. The same signalling molecule can act on different cell types as long as the right type of receptors exist on the targets. (e.g. acetylcoline acting on heart muscle and skeletal muscle cells: both cell types have receptors that bind to acetylcoline, but their downstream machinery is different so the response is different)
How is signalling specificity achieved within the cells?
There are numerous pathways interacting with one another inside the cells, so what prevents the upstream signals from activating all the possible downstream targets?
1) Preformed signalling complexes on a scaffold protein:Scaffold proteins assemble the target intracellular signalling proteins ahead of time, and when a signal molecule activates the receptor, the activated local complex is forced to work together to activate downstream signals even if they might be able to activate other targets had they been separated.
* the activation pattern is forced to be: 1 —> 2—> 3 because of their proximity
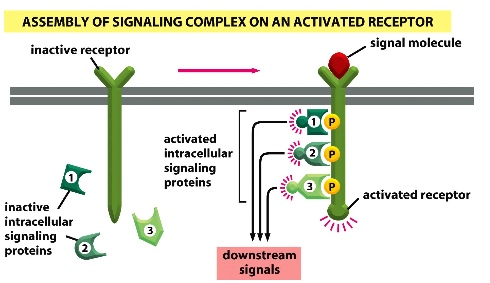
2) Assembly of signalling complex on an activated receptor: Signal binding to receptor induces the assemble of intracellular signalling proteins onto the activated receptors —> downstream signals
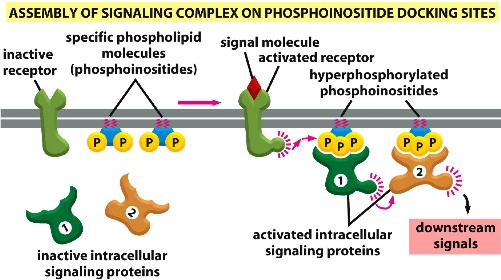
3) Assembly of signalling complex on phosphoinositide (PIP) docking sites: signal-receptor complex activates local PIPs, PIPs can bring together specific proteins —> downstream signals
How are signalling pathways coordinated?
Combination of different signals can lead to multiple outcomes
Coincidence detectors only activate downstream signals when two upstream signals are both detected. —> One signal phosphorylates one side, another one phosphorylates another.
Organization in space:
primary cilia: some key signalling pathways are found inside these: —> insulating the pathway from the rest of the cell
synapses: insulating the signalling complex
Organization in time:
synaptic signalling: very fast
endocrine signally: relatively slow due to the speed of blood flow
response speed to a signal can be slow or fast depending on the availability of cellular machinery: whether or not it has been built.
Feedback mechanism in signalling pathways
positive or negative —> downstream pathway activated + promotion or inhibition of the loop
positive feedback leads to a long lasting signal, and if it’s strong enough, can become self sustaining so that the system is kept at high activation even when the signal is lost.
e.g. when cells differentiate, signal F and G contributes to the self-sustaining of differentiation
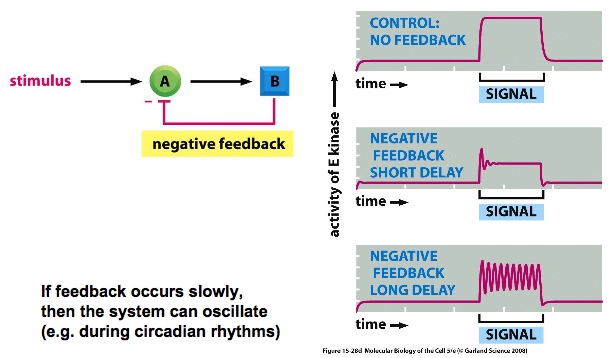
negative feedback: duration of the signal is maintained. If feedback occurs quickly, signalling is suppressed. Allows cells to respond to a range of signal strengths
system can adapt to the activation so there needs to be more activation to boost signal.
slow negative feedback can make the system oscillate. —> the delay from signalling to inhibiting signalling creates the pattern.